|
edit SideBar
|
Tertiary
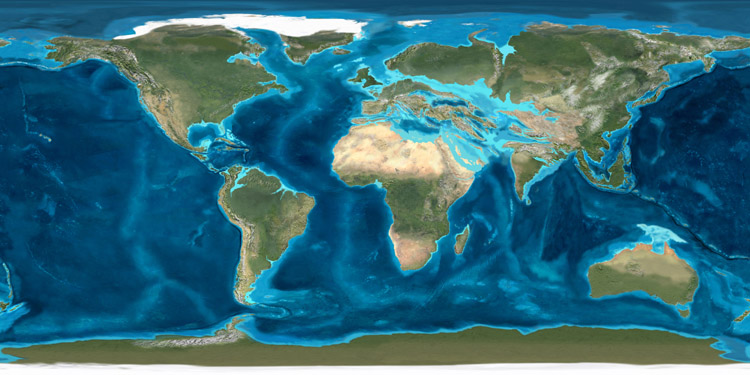
Map of the Oligocene (35 ma)
Copyright by Ron Blakey
During the Oligocene Australia and South America were completely separated from Antarctica. The Separation of South America from Antarctica led to an Antarctic circumpolar current, which consequence is a cooling down of Antarctica, followed by creation of an ice shield, resulting in a worldwide marine transgression.
India collided with Asia.
The cooling resulted in a fragmentation of biotic provinces and a reduced diversity of marin plancton, which is a starting point of the food chain.
The Oligocene is divided into the epochs:
2008
L. Alegret, et al. 2008, p. 94:
- "The Oligocene in particular is considered a period marked by large and abrupt climate changes, paleogeographic changes including the opening of the Tasmanian Gateway and Drake Passage, large fluctuations in the volume of the Antarctic Ice Sheet after its initiation in the earliest Oligocene, with related eustatic changes at orbital frequencies (e.g., Wade and Pälike, 2004; Pälike et al., 2006). The formation of cold deep water in the Southern Ocean and/or in the northern Atlantic may have started in the earliest Oligocene (Kennett, 1977; Lawyer and Gahagan, 2003; Miller et al., 1991, 2005; Zachos et al., 2001), but the timing and pattern of these circulation changes is under vigorous debate (e.g., Barker and Thomas, 2004; Scher and Martin, 2004; Via and Thomas, 2006; Thomas and Via, 2007; Thomas et al., 2008). This transitional period in Earth's history was characterised by strong, short-term fluctuations between warmer and colder intervals that have been recognised and at least tentatively correlated around the world (Zachos et al., 2001). These climate fluctuations occur at orbital frequencies (Wade and Pälike, 2004; Coxall et al., 2005; Pälike et al., 2006), with some of the more extreme cold events (formerly called Oievents) occurring at low obliquity."
Rasser, Harzhauser et al. 2008, p. 1040:
- "The Oligocene began with a major transgression during NP21 which flooded the Egeln-Halle Lignite District, the Leipzig district (Standke 1997), and large parts of the German Mittelgegirge (Ritzkowski 1999b). After short regressive phase (Vandenberghe et al. 2002) a minor transgression was observed in the NSB [North Sea Basin] in zone NP22 (Gürs 2005), which flooded the Upper Rhine Valley via the Hessian Depression (Martini 1973). The most striking transgression took place in the early NP23 when the sea spread over vast areas of the low mountain ranges (Ritzkowski 1987) flooding the Leipzig Bay (Müller 1983; Eissmann & Litt 1994), the Upper Rhine Valley, the Lower Rhine Embayment and northern Belgium (Vandenberghe et al. 2002). Neogene development of the NSB is already evident in the Late Oligocene following a severe regressive phase when the basin was clearly delimited to the east and river systems brought in vast amounts of sediment from the NE and east. Greater parts of the Fennoscandinavian Shield were lowlands and low mountains. During the Chattian transgression, marine conditions were restored in large parts of the NSB. The NE part of Denmark and the Lower Rhine District were slowly covered by river systems and the coastline retreated inwards (Dybkjaer 2004a). Northern parts of eastern Germany were marine influenced throughout the entire Late Oligocene (Lotsch 1981; Standke 2001, 2008a). In the SW a semicontinous connection to the Atlantic developed as shown by faunal evidence (R. Janssen 1979). Other pathways, such as the northern one around Scotland, have been discussed as the only connection (Gripp 1958) but single tropical molluscs such as Morum, Perrona and Mitrolumna raulini septentrionalis Janssen 1979 require a southern connection, the latter showing an affinity to the Chattian of the Aquitaine Basin. A warm climate led to the invasion of a nearly subtropical marine fauna. It is preserved in the Voort Sands of Northern Belgium, the Grafenberg Sands of the Lower Rhine Embayment, the Kassel Sands (showing a strong ingression into the Hessian Depression in the early Late Oligocene) and the famous Sternberg erratic boulders of Mecklenburg (Bülow & Müller 2004) and Glaukonitsand in the Lusatia area (Standke 2006). Late Oligocene sediments in northern Belgium are restricted to the Roermond Graben and a small portion of the Central Graben area. Deep water conditions prevailed in the basin centre. In the Late Chattian a coast with lagoons and barrier islands developed in the middle Jutland and this was related to the Ringkobing-Fyn High (Dybkjaer 2004a, b).
2014
Garilli & Parrinello, 2014, p. 401-403:
- "The Late Eocene–Oligocene palaeogeographic reconstructions of those areas (Vincent 1990; Rögl 1999: figs. 2, 3; Rasser et al. 2008, with references) outline the North Sea and Paris Basins (N Atlantic realm) as distinct marine provinces separated by the Thule bridge, a wide land-passageway putting Great Britain in communication with central-western continental Europe until the Late Oligocene. At this time sporadic communications between the Atlantic and North Sea Basin occurred in the Channel area, as indicated by Janssen (1978b) on the basis of some Late Oligocene marine faunal affinities between the North Sea Basin and the Aquitaine Basin. However, it is possible that a southernmost connection favoured these interchanges as supposed by Rasser et al. (2008). The distribution of G. biangulata (Fig. 17) indicates that a faunal interchange (possibly unidirectional) between the North Sea and Paris Basins started in the Early Oligocene, when the Thule land-bridge was already in-filled (at least intermittently) in the Channel area, much earlier than had been supposed.
- [...] Notwithstanding this, Galeodinopsis seems never to have lived in the eastern part of the southern margin of the North Sea Basin, nor in the Paratethyan area, even though communication between the North Sea and Mediterranean/Paratethyan areas was established during that time, especially in the Early Oligocene, when ocean circulation supplied water from the North Sea to the Paratethys (Rögl 1999: figs. 1, 2). The distributional gap in the North Sea Basin may be at least partially explained by the paralic condition that affected the southeastern part of the North Sea Basin during Late Oligocene time (Rasser et al. 2008).
- [...] Tropical to warm-temperate conditions have been inferred for the Palaeogene of Europe (Vincent 1990) and, more specifically, subtropical conditions were indicated for the Late Oligocene of the North Sea Basin (Rasser et al. 2008). Actually, a warm (possibly tropical-subtropical) climatic regime in the Paleogene of the North Sea Basin is suggested by the analogies (at least at genus level) between the highly diverse molluscan assemblages from the Mediterranean Neogene and the Late Eocene–Oligocene of Germany, southwards to the Mainz basin: several molluscan taxa characteristic for warm climate, such as Bathytoma Harris and Burrows, 1891, Isognomon Lightfoot, 1786, Ficus Röding, 1798, Gemmula Weinkauff, 1875 and others, have been recorded in those areas (Janssen 1978b, 1979; Gründel 1997; Monegatti and Raffi 2001; Müller 2011). Furthermore, warm, tropical to subtropical, marine climatic conditions were inferred for this part of central Europe with particular regard to the Latdorfian faunal complex and similar assemblages of younger (Oligocene) age (Müller 2011). It is remarkable that from the middle Eocene onwards, the Cenozoic continental history of central Europe, in accordance with global marine oxygen isotope records, was characterized by the increasing degree of seasonality, and a short-term cooling at the base of the Aquitanian, approximating the Oligocene/Miocene boundary, was recorded in the Weisselster Basin, in central Germany (Mosbrugger et al. 2005). Also from a palaeogeographical point of view, the area of central-western Europe underwent remarkable changes. During the Early Miocene, marine regressive conditions rewrote the palaeogeography of the North Sea Basin, triggering the progressive reduction of its southernmost part, which was constrained to higher (presumably relatively colder) latitudes. Regressive conditions also caused the disappearance of satellite basins such as those in the Rhine Graben (e.g., the Mainz Basin; Rögl 1999). This palaeogeographic-climatic evolution was possibly responsible for the disappearance of Galeodinopsis from the mid-high latitudes of Central Europe and for its migration to the Iberian Atlantic coasts and into the (paleo)Mediterranean during Mio-Pliocene time. At the end of the Pliocene, at around 2.6 My B.P. (approximating the Piacenzian/Gelasian boundary), the marked onset of the continental ice sheet in the Northern Hemisphere (Shackleton et al. 1984) triggered stepwise climatic deteriorations at lower latitudes, causing the Mediterranean extinction of several molluscan taxa (Monegatti and Raffi 2001). The new climatic regime probably was the main cause of the disappearance of Galeodinopsis from the Mediterranean and its further migration towards the subtropical-tropical West Africa coasts, where this genus still lives. As for the ecological/bathymetrical requirements of Galeodinopsis, it is remarkable that for the Late Paleogene around Magderburg and Glimmerode (southern part of the North Sea Basin), where G. biangulata and G. semperi lived, the palaeofauna indicates normal marine conditions in rocky-sandy shore environments (Janssen 1978b; Mueller 2008, 2011). Also, the requirements of G. tiberiana are comparable, as indicated by its occurrence in the Pliocene sands of the Mediterranean Altavilla site, the depositional setting of which was mainly referred to infra-circalittoral shelf conditions (Dell’Angelo et al. 2012)."
References
- Akbar-Baskalayeh, N., Lees, G., Ghasemi-Nejad, E., Yazdi-Moghadam, M., Pignatti J. (2020) Biometric study of late Oligocene larger benthic Foraminifera (Lepidocyclinidae and Nummulitidae) from the Qom Formation, Central Iran (Tajar-Kuh section). Journal of Paleontology, page 1 of 23
- L. Alegret, L.E. Cruz, R. Fenero, E. Molina, S. Ortiz & E. Thomas, 2008. Effects of the Oligocene climatic events on the foraminiferal record from Fuente Caldera section (Spain, western Tethys); Palaeogeography, Palaeoclimatology, Palaeoecology 269 (2008): 94–102.
- A. Azzaroli. 1958. L'Oligocene e il Miocene della Somalia. Stratigrafia, Tettonica, Paleontologia (Macroforaminiferi, Coralli, Molluschi). Palaeontolographia Italica 52:1-142.
- Batjes, D.A.J. (1958). Foraminifera of the Oligocene of Belgium. Memoirs of the Royal Belgian Institute of Natural Sciences, first series, 143, 1958
- Benedetti, A. (2017). Eocene/Oligocene deep-water agglutinated foraminifers (DWAF) assemblages from the Madonie Mountains (Sicily, Southern Italy). Palaeontologia Electronica, 20.1, 2017
- Bosellini F. & Perrin C. 2008. Estimating Mediterranean Oligocene-Miocene sea-surface temperatures: An approach based on coral taxonomic richness. Palćogeography, Palćoclimatology, Palćoecology, Amsterdam, vol. 258, p. 71-88.
- Cadée, M.C. 1990. Het Oligoceen en de Oligocene Mollusken Fauna's van Nederland en omgeving; talk 10. Nov. 1990 in Amsterdam, Fulltext
- Cossmann M. (1892).- Révision sommaire de la faune du terrain oligocčne marin aux environs d'Étampes.- Journal de Conchyliologie, Paris, t. 40, p. 330-375.
- Cossmann M. (1893).- Révision sommaire de la faune du terrain oligocčne marin aux environs d'Étampes (suite).- Journal de Conchyliologie, Paris, t. 41, p. 297-363.
- Dockery D.T. III & Lozouet P. (2006).- Eocene-Oligocene molluscan extinctions and first appearance data across the North Atlantic in the North American Gulf Coastal Plain and Europe. In: Prothero D.R., Ivany L.C. & Nesbitt E.A. (eds.), From Greenhouse to Icehouse. The marine Eocene-Oligocene transition.- New York, Columbia University Press, p. 303-340.
- D. Esu, O. Girotti & J. Pignatti, 2005. Late Oligocene-?Miocene mollusc and foraminiferal assemblages from the vicinity of Otranto (Southern Apulia, Italy): a non-marine to marine transition; Rendiconti Soc. Paleont. Ital., 2(2005):75-85
- Garilli, V. and Parrinello, D. 2014. Taxonomy and palaeobiogeography of the Cenozoic Euro-Mediterranean rissoid gastropod Galeodinopsis and its relationship with close genera. Acta Palaeontologica Polonica 59 (2): 379–406, Fulltext
- Glibert, M. 1957. Pélécypodes et gastropodes du Rupélien supérieur et du Chattien de la Belgique; Mémoires du Musée royal d'Histoire naturelle de Belgique 137:1-98
- Glibert, M. & De Heinzelin, J. 1954. L'Oligocčne inférieur belge. Volume Jubilaire Victor Van Straelen I, Brussel, IRScNB. pp. 281-438.
- Gürs K. (1995, unpublished).- Revision der marinen Molluskenfauna des Unteren Meeressandes (Oligozän, Rupelium) des Mainzer Beckens.- Dissertation, München, 314 p. (64 Pls.).
- Gürs K. & Moths H. (2002).- Neues zu den Rissooidea (Mollusca, Gastropoda) aus dem Rupelium (Oligozän) des Mainzer Beckens und des Nordseebeckens.- Courier Forschungsinstitut Senckenberg, Frankfurt, vol. 237, p. 255-273.
- Harder, P. 1913. De oligocaene Lag i Jaernbanegennemskaeringen ved Arhus Station; Danmarks geologiske Undersoegelse (II) 22, pp. 1-140. Copenhagen
- Harzhauser M. (2004).- Oligocene gastropod Faunas of the Eastern Mediterranean (Mesohellenic Trough/Greece and Esfahan-Sirjan Basin/Central Iran).- Courier Forschungsinstitut Senckenberg, Frankfurt, vol. 248, p. 93-181.
- Harzhauser M., Piller W.E. & Steininger F.E. (2002).- Circum-Mediterranean Oligo-Miocene biogeographic evolution - the gastropods' point of view.- Palćogeography, Palćoclimatology, Palćoecology, Amsterdam, vol. 183, p. 103-133.
- Harzhauser M., Kroh A, Mandic O., Piller W.E., Göhlich U., Reuter M. & Berning B. (2007).- Biogeographic responses to geodynamics: A key study all around the Oligo-Miocene Tethyan Seaway.- Zoologischer Anzeiger, Berlin, vol. 246, nş 4, p. 241-256.
- Janssen A.W. (1984b).- Late Oligocene molluscs from a sand-pit near Máriahalom (Hungary): A preliminary study.- Annales Universitatis Scientiarum Budapestinensis de Rolando Eötvös Nominatae Sectio Geologica, Budapest, vol. 24, p. 111-149 (4 Pls.).
- Janssen, A.W., 1989. Some new pteropod species from the North Sea Basin Cainozoic (Mollusca: Gastropoda, Euthecosomata). Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie 26 (3): 91-133
- Janssen R. (1978a).- Die Scaphopoden und Gastropoden des Kasseler Meeressandes von Glimmerode (Niederhessen).- Geologisches Jahrburch, Hannover, (Reihe A), Band A 41, 195 p. (7 Pls.).
- Janssen R. (1978b).- Die Mollusken des Oberoligozäns (Chattium) im Nordsee-Becken, 1 - Scaphopoda, Archaeogastropoda, Mesogastropoda.- Archiv für Molluskenkunde, Frankfurt, vol. 109, nş 1-3, p. 137-227 (Pls. 9-14).
- Janssen R. (1979).- Die Mollusken des Oberoligozäns (Chattium) im Nordsee-Becken. 2. Neogastropoda, Euthyneura, Cephalopoda.- Archiv für Molluskenkunde, Frankfurt, vol. 109, nş 4, p. 277-376 (5 Pls.).
- Lozouet P. (coord., 2012). Stratotype Stampien. Biotope, Mčze; Patrimoine géologique, Muséum national d'Histoire naturelle, Paris, vol. 4, 464 p. (1 CD-Rom).
- Pierre Lozouet, 2014. Temporal and latitudinal trends in the biodiversity of European Atlantic Cenozoic gastropod (Mollusca) faunas. A base for the history of biogeographic provinces; Carnets de Géologie [Notebooks on Geology] vol. 14 (14), p. 273-314, Fulltext
- Maréchal, R. 1991. A new lithostratigraphic scale for the Paleogene of Belgium. Abstracts 3rd Biannual Joint Meeting Regional Committees on Northern Palaeogene and Northern Neogene Stratigraphy, Gent, 9-13 September 1991, 2 p., 1 fig.
- Maréchal, R. & Laga, P., 1988. Voorstel Lithostratigrafische Indeling van het Paleogeen. Nationale Commissies voor Stratigrafie, Commissie Tertiair, Belgische Geologische Dienst, Brussel, 208 pp.
- Moths, H., 2000. Die Molluskenfauna im Rupelton der Ziegeleitongrube Malliss im Wanzeberg (südwestl. Mecklenburg-Vorpommern). Malliss, Regionalmuseum. 103 pp.
- A. Müller, 2013. Exkursionsführer zur Sitzung der Subkommission Tertiär-Stratigraphie 20-22. März 2013 in Leipzig. Subkommission Tertiär-Stratigraphie (SKT), Fulltext
- Premoli Silva, I. & Jenkens, D.G., 1993. Decision on the Eocene-Oligocene boundary stratotype. Episodes, 16(3): 379-382.
- Ramos, E. & Martinell, J. 1985. Datos preliminares sobre la malacofauna marina del Oligoceno de Mallorca; Iberus 5:1-9, Fulltext
- Rasser, M.W. & Harzhauser, M. (co-ordinators) et al. 2008. Palaeogene and Neogene in Tom McCann (Ed.), The Geology of Central Europe, Vol. 2, The Geology of Central Europe: Mesozoic and Cenozoic, Geological Society of London, 2008, p. 1031-1139.
- Schnetler, K. I. & Beyer, C. 1990. A Late Oligocene (Chattian B) mollusc fauna from the coastal cliff at Mogenstrup, North of Skive, Jutland, Denmark. Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie (Contributions to Tertiary and Quaternary Geology) 27 (2-3), 39-81, Fulltext
- Schnetler, K.I. & Palm, E., 2008: The molluscan fauna of the Late Oligocene Branden Clay/Denmark. – Palaeontos 15, 1–92. Mortsel.
- Vandenberghe, N., 1978. Sedimentology of the Boom Clay Formation, Belgium. Bulletin van de Belgische Vereniging voor Geologie 102 (1-2): 5-77.
- Vernyhorova, Y.V. & Ryabokon, T.S.(2020). The stratigraphy of the Oligocene-lower Miocene deposits of southern Ukraine. Turkish Journal of Earth Sciences, (2020), 29; 170-207
- Yazdi-Moghadam M. (2011) - Early Oligocene Larger Foraminiferal Biostratigraphy of the Qom Formation, South of Uromieh (NW Iran)
Internet
|